Herança do Sexo II

Herança do Sexo II
Herança Restrita ao Sexo
Dizemos que uma herança é restrita ao sexo quando está relacionada ao cromossomo Y, na sua porção não homóloga ao cromossomo X. É restrita porque só ocorre em homens, pois as mulheres não possuem cromossomo Y.
Os genes localizados apenas no cromossomo Y são chamados genes holândricos, do grego holos = todo; andros = masculino.
Herança influenciada pelo sexo
Ocorre quando a doença ou característica se manifesta de forma diferente em homens e mulheres.
A calvície é condicionada por um gene dominante. Nas mulheres, ele se expressa apenas em dose dupla.

Isso ocorre porque o gene C atua como dominante na presença do hormônio masculino testosterona, e pode se expressar mesmo em dose simples. Como as mulheres possuem baixíssimas quantidades desse hormônio, o gene em dose simples não é suficiente para se expressar.
Alguns casos de anomalias humanas
Consideramos, a seguir, três casos em que a herança sexual pode determinar anormalidade na espécie humana.Síndrome de Turner

Os indivíduos portadores dessa síndrome não tem um dos cromossomos x, apresentando um cariótipo igual a 44A + XO.
Exibem, entre outras características, fenótipo feminino, ovários atrofiados e estatura baixa.
Síndrome de Klinefelter

Neste caso os indivíduos possuem um cromossomo X extra. Manifesta-se apenas no homem, cujo cariótipo será 44A + XXY.
O individuo apresenta um fenótipo masculino, testículos atrofiados, deficiência mental e pequenos seios, entre outras características.
Síndrome de Down

Os indivíduos afetados apresentam um autossomo extra. Essa anomalia pode ocorrer tanto nos homens como nas mulheres, cujos cariótipos serão, respectivamente, 45A + XY e 45A + XX.
Os portadores da síndrome de down apresentam, entre outras características, deficiência mental, tendencia a leucemia e mãos curtas e largas.
Trata-se de umas das anomalias mais frequentes. É também conhecida como trissomia do cromossomo 21, isto é, em vez de dois cromossomos, o par de 21 de cromossomos autossômicos apresenta três cromossomos.
Aneuploidias: o cariótipo alterado
{kind=link}
Os fenômenos em que ocorre variação numérica de um ou mais cromossomos no cariótipo do individuo são chamados aneuploidias. Resultam de uma não-disjunção, isto é, de uma não-separação de certos pares de homólogos durante a formação de gametas.
Na perda de um cromossomo, a aneuploidia é do tipo monossômica (representando = 2n - 1), como ocorre na síndrome de Turner.
No acréscimo de um cromossomo tem-se a trissomia ( representação = 2n + 1), como ocorre nas síndromes de Klinefelter e de Down.
Herança do Sexo I
Na espécie humana, as células somáticas contem 46 cromossomos ou 23 pares de homólogos. Destes, 23 são de origem paterna e 23 de origem materna. No entanto, a determinação do sexo do individuo resulta da interação de determinados genes situados num único par de homólogos. esse par é formado pelos chamados cromossomos sexuais, conhecidos também como heterossomos ou alossomas.
Os demais cromossomos constituem os autossomos, que não tem implicação com o sexo.
 Cromossomos sexuais femininos e masculinos
Cromossomos sexuais femininos e masculinos
Os dois cromossomos sexuais existentes nas mulheres exibem mesma forma e são denominados cromossomos x. Nos homens, porém, existe apenas um cromossomo x; o outro, de forma recurvada, é denominado cromossomo y. Assim, temos:
- Homens:
44 autossomos + 2 sexuais
44A+ XY
- Mulheres
44A+ XX
Sexo feminino: Homogamético
Pelo que sabemos sobre a distribuição cromossômica, durante o processo de formação dos gametas, uma mulher produz apenas um tipo de gameta; daí o sexo feminino ser homogamético:
♀ 44A + XX → ovulogênese → 22A + X → óvulo
Sexo Masculino: Heterogamético
Um homem, por sua vez, produz dou tipos básicos de gametas, sendo, por isso, considerado heterogamético:
♂ = 44A + XY → espermatogênese → 22A + X
♂= 44A + XY → espermatogênese→ 22A + Y
As drosófilas ( moscas-das-frutas) apresentam o mesmo tipo de determinação cromossômica para o sexo que é encontrado na espécie humana. Já entre as aves o sexo heterogamético é o feminino sendo sexo masculino homogamético. Assim, chamado de X e Y os cromossomos sexuais da drosófila e de Z e W os das aves, temos a seguinte sempre situações:
Drosófilas Aves
♂= 2A + XY ♂= 2A + ZZ
♀= 2A + XX ♀= 2A + ZW
Herança ligada ao sexo
A herança é considerada ligada ao sexo quando os genes envolvidos situam-se no cromossomo X, em sua porção não-homóloga,isto é, sem correspondência no cromossomo Y.
A herança ligada ao sexo refere-se aos genes localizados em cromossomos sexuais que estão envolvidos na determinação de características.
Na espécie humana, o cromossomo sexual masculino Y apresenta poucos genes. Já, o cromossomo sexual feminino X possui grande quantidade de genes envolvidos na determinação de várias características.
Os cromossomos XY apresentam pequenas regiões homólogas em suas extremidades. Assim, praticamente não há recombinação entre os seus genes.
Os genes localizados no cromossomo X que têm alelo correspondente no cromossomo Y, seguem o padrão da herança ligada ao sexo.
Assim, a herança ligada ao sexo está restrita aos cromossomos sexuais. Enquanto, a herança autossômica é a que ocorre nos cromossomos autossômicos.
Estudaremos a seguir duas anomalias cuja herança está ligada ao sexo: daltonismo e hemofilia.
Hemofilia

É um distúrbio da coagulação sanguínea, em que falta o fator VIII, uma das proteínas envolvidas no processo, encontrado no plasma das pessoas normais. As pessoas hemofílicas têm uma tendência a apresentarem hemorragias graves depois de traumatismos banais, como um pequeno ferimento ou uma extração dentária.O tratamento da hemofilia consiste na administração do fator VIII purificado ou de derivados de sangue em que ele pode ser encontrado (transfusões de sangue ou de plasma). Pelo uso frequente de sangue e de derivados, os pacientes hemofílicos apresentam uma elevada incidência de AIDS e de hepatite tipo B, doenças transmitidas através dessas vias.

A hemofilia atinge cerca de 300.000 pessoas. É condicionada por um gene recessivo, representado por h, localizado no cromossomo X. É pouco freqüente o nascimento de mulheres hemofílicas, já que a mulher, para apresentar a doença , deve ser descendente de um hímen doente (XhY) e de uma mulher portadora (XHXh) ou hemofílica (XhXh). Como esse tipo de cruzamento é extremamente raro, acredita-se que praticamente inexistiriam mulheres hemofílicas. No entanto, já foram relatados casos de hemofílicas, contrariando assim a noção popular de que essas mulheres morreriam por hemorragia após a primeira menstruação (a interrupção do fluxo menstrual deve-se à contração dos vasos sanguíneos do endométrio, e não a coagulação do sangue).
Daltonismo

Trata-se da incapacidade relativa na distinção de certas cores que, na sua forma clássica, geralmente cria confusão entre o verde e o vermelho.
É um distúrbio causado por um gene recessivo localizado na porção heteróloga do cromossomo X, o gene Xd, enquanto o seu alelo dominante XD determina a visão normal.

A mulher de genótipo XDXd, embora possua um gene para o daltonismo, não manifesta a doença, pois se trata de um gene recessivo. Ela é chamada de portadora do gene para o daltonismo. O homem de genótipo XdY, apesar de ter o gene Xdem dose simples, manifesta a doença pela ausência do alelo dominante capaz de impedir a expressão do gene recessivo.
Homem XdY não é nem homozigoto ou heterozigoto: é hemizigoto recessivo, pois do par de genes ele só possui um. O homem de genótipo XDY é hemizigoto dominante.

Se você consegue distinguir perfeitamente o número 74 entre as bolinhas da figura acima,
então você não é daltônico.
|
Grupos Sanguíneos: Fator Rh; sistema MN

Fator Rhesus ( fator Rh)
Um terceiro sistema de grupos sanguíneos foi descoberto a partir dos experimentos desenvolvidos por Landsteiner e Wiener, em 1940, com sangue de macaco do gênero Rhesus. Esses pesquisadores verificaram que ao se injetar o sangue desse macaco em cobaias, havia produção de anticorpos para combater as hemácias introduzidas.
Ao centrifugar o sangue das cobaias obteve-se o soro que continha anticorpos anti-Rh e que poderia aglutinar as hemácias do macaco Rhesus. As conclusões daí obtidas levariam a descoberta de um antígeno de membrana que foi denominado Rh (Rhesus), que existia nesta espécie e não em outras como as de cobaia e, portanto, estimulavam a produção anticorpos, denominados anti-Rh.
Há neste momento uma inferência evolutiva: se as proteínas que existem nas hemácias de vários animais podem se assemelhar isto pode ser um indício de evolução. Na espécie humana, por exemplo, temos vários tipos de sistemas sanguíneos e que podem ser observados em outras espécies principalmente de macacos superiores.
Analisando o sangue de muitos indivíduos da espécie humana, Landsteiner verificou que, ao misturar gotas de sangue dos indivíduos com o soro contendo anti-Rh, cerca de 85% dos indivíduos apresentavam aglutinação (e pertenciam a raça branca) e 15% não apresentavam. Definiu-se, assim, "o grupo sanguíneo Rh+ ” ( apresentavam o antígeno Rh), e "o grupo Rh- " (não apresentavam o antígeno Rh).
No plasma não ocorre naturalmente o anticorpo anti-Rh, de modo semelhante ao que acontece no sistema Mn. O anticorpo, no entanto, pode ser formado se uma pessoa do grupo Rh -, recebe sangue de uma pessoa do grupo Rh +. Esse problema nas transfusões de sangue não são tão graves, a não ser que as transfusões ocorram repetidas vezes, como também é o caso do sistema MN.
A Herança do Sistema Rh
Três pares de genes estão envolvidos na herança do fator Rh, tratando-se portanto, de casos de alelos múltiplos.
 Para simplificar, no entanto, considera-se o envolvimento de apenas um desses pares na produção do fator Rh, motivo pelo qual passa a ser considerado um caso de herança mendeliana simples. O gene R, dominante, determina a presença do fator Rh, enquanto o gene r, recessivo, condiciona a ausência do referido fator. Para simplificar, no entanto, considera-se o envolvimento de apenas um desses pares na produção do fator Rh, motivo pelo qual passa a ser considerado um caso de herança mendeliana simples. O gene R, dominante, determina a presença do fator Rh, enquanto o gene r, recessivo, condiciona a ausência do referido fator.Doença hemolítica do recém-nascido ou eritroblastose fetal
Uma doença provocada pelo fator Rh é a eritroblastose fetal ou doença hemolítica do recém-nascido, caracterizada pela destruição das hemácias do feto ou do recém-nascido. As conseqüências desta doença são graves, podendo levar a criança à morte.
Durante a gestação ocorre passagem, através da placenta, apenas de plasma da mãe para o filho e vice-versa devido à chamada barreira hemato-placentária. Pode ocorrer, entretanto, acidentes vasculares na placenta, o que permite a passagem de hemácias do feto para a circulação materna. Nos casos em que o feto possui sangue fator rh positivo os antígenos existentes em suas hemácias estimularão o sistema imune materno a produzir anticorpos anti-Rh que ficarão no plasma materno e podem, por serem da classe IgG, passar pela BHP provocando lise nas hemácias fetais. A produção de anticorpos obedece a uma cascata de eventos (ver imunidade humoral) e por isto a produção de anticorpos é lenta e a quantidade pequena num primeiro. A partir da segunda gestação, ou após a sensibilização por transfusão sanguínea, se o filho é Rh + novamente, o organismo materno já conterá anticorpos para aquele antígeno e o feto poderá desenvolver a DHPN ou eritroblastose fetal.
O diagnóstico pode ser feito pela tipagem sanguínea da mãe e do pai precocemente e durante a gestação o teste de Coombs que utiliza anti-anticorpo humano pode detectar se esta havendo a produção de anticorpos pela mãe e providências podem ser tomadas. Uma transfusão , recebendo sangue Rh -, pode ser feita até mesmo intra-útero já que Goiânia está se tornando referência em fertilização in vitro. O sangue Rh - não possui hemácias com fator Rh e não podem ser reconhecidas como estranhas e destruídas pelos anticorpos recebidos da mãe. Após cerca de 120 dias, as hemácias serão substituídas por outras produzidas pelo próprio indivíduo. O sangue novamente será do tipo Rh +, mas o feto já não correrá mais perigo

Após o nascimento da criança toma-se medida profilática injetando, na mãe Rh- , soro contendo anti Rh. A aplicação logo após o parto, destrói as hemácias fetais que possam ter passado pela placenta no nascimento ou antes. Evita-se , assim, a produção de anticorpos “zerando o placar de contagem”. Cada vez que um concepto nascer e for Rh+ deve-se fazer nova aplicação pois novos anticorpos serão formados.
Os sintomas no RN que podem ser observados são anemia (devida à destruição de hemácias pelos anticorpos), icterícia (a destruição de hemácias aumentada levará a produção maior de bilirrubina indireta que não pode ser convertida no fígado), e após sua persistência o aparecimento de uma doença chamada Kernicterus que corresponde ao depósito de bilirrubina nos núcleos da base cerebrais o que gerará retardo no RN.
Grupos sanguíneos do Sistema MN No sistema MN de grupos sanguíneos existem três grupos diferentes. O grupo M, no qual pertencem as pessoas que possuem o antígeno M em suas hemácias. O grupo N, cujas pessoas possuem o antígeno Nem suas hemácias, e o grupo MN, no qual as pessoas possuem o antígeno M e o antígeno N em suas hemácias, lembrando que esses grupos sanguíneos são determinados por um par de alelos sem relação de dominância entre si. No sistema MN de grupos sanguíneos existem três grupos diferentes. O grupo M, no qual pertencem as pessoas que possuem o antígeno M em suas hemácias. O grupo N, cujas pessoas possuem o antígeno Nem suas hemácias, e o grupo MN, no qual as pessoas possuem o antígeno M e o antígeno N em suas hemácias, lembrando que esses grupos sanguíneos são determinados por um par de alelos sem relação de dominância entre si.
A importância desse sistema em transfusões sanguíneas não é relevante, pois no plasma sanguíneo não encontramos anticorpos (aglutininas) que combatem esse tipo de antígeno, sendo que eles são produzidos somente quando recebem algum tipo de estímulo, por exemplo, quando uma pessoa do grupo M recebe sangue de outra pessoa do grupo N. Mesmo quando o corpo da pessoa recebe esse estímulo, ela produz uma quantidade muito pequena de anticorpos, não causando nenhum tipo de prejuízo à pessoa.
Como sabemos que o sistema MN de grupos sanguíneos é um caso de ausência de dominância entre dois alelos, podemos definir seus genótipos da seguinte forma:

A letra L utilizada é uma homenagem aos pesquisadores Landsteiner e Levine.
Transfusões no Sistema MN
A produção de anticorpos anti-M ou anti-N ocorre somente após sensibilização (você verá isso no sistema RH). Assim, não haverá reação de incompatibilidade se uma pessoa que pertence ao grupo M, por exemplo, receber o sangue tipo N, a não ser que ela esteja sensibilizada por transfusões anteriores.
|
Grupos sanguíneos : Sistema ABO (polialelia: alelos múltiplos)
 Neste Tema vamos estudar que há casos, em que um determinado caráter pode ser condicionado por três ou mais genes alelos contrastantes: trata-se da polialelia, ou seja, da ocorrência de uma série de alelos múltiplos.
Neste Tema vamos estudar que há casos, em que um determinado caráter pode ser condicionado por três ou mais genes alelos contrastantes: trata-se da polialelia, ou seja, da ocorrência de uma série de alelos múltiplos.
Embora na polialelia a espécie possa exibir três ou mais formas alternativas de um mesmo gene, em um determinado indivíduo existirão apenas dois genes alelos, sendo um de origem materna e outra de origem paterna. isso se justifica, uma vez que as células somáticas ( diploides, 2n) contém cromossomo homólogos distribuídos aos pares, isto é, combinados dois a dois; como em cada lócus existe apenas um gene, conclui-se que na determinação de um caráter os alelos interagem dois a dois.
Como por exemplo de polialelia consideramos os grupos sanguíneos humanos do sistema ABO.
A descoberta dos grupos sanguíneos
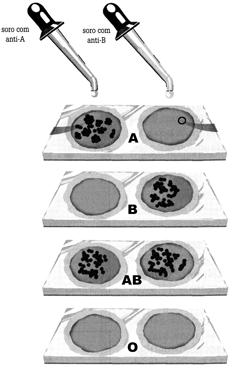 |
| Determinação dos grupos sanguíneos utilizando soros anti-A e anti-B. Amostra 1- sangue tipo A. Amostra 2 - sangue tipo B. Amostra 3 - sangue tipo AB. Amostra 4 - sangue tipo O. |
Por volta de 1900, o médico austríaco Karl Landsteiner (1868 – 1943) verificou que, quando amostras de sangue de determinadas pessoas eram misturadas, as hemácias se juntavam, formando aglomerados semelhantes a coágulos. Landsteiner concluiu que determinadas pessoas têm sangues incompatíveis, e, de fato, as pesquisas posteriores revelaram a existência de diversos tipos sanguíneos, nos diferentes indivíduos da população.
Quando, em uma transfusão, uma pessoa recebe um tipo de sangue incompatível com o seu, as hemácias transferidas vão se aglutinando assim que penetram na circulação, formando aglomerados compactos que podem obstruir os capilares, prejudicando a circulação do sangue.
Aglutinogênios e aglutininas
No sistema ABO existem quatro tipos de sangues: A, B, AB e O. Esses tipos são caracterizados pela presença ou não de certas substâncias na membrana das hemácias, os aglutinogênios, e pela presença ou ausência de outras substâncias, as aglutininas, no plasma sanguíneo.
Existem dois tipos de aglutinogênio, A e B, e dois tipos de aglutinina, anti-A e anti-B. Pessoas do grupo A possuem aglutinogênio A, nas hemácias e aglutinina anti-B no plasma; as do grupo B têm aglutinogênio B nas hemácias e aglutinina anti-A no plasma; pessoas do grupo AB têm aglutinogênios A e B nas hemácias e nenhuma aglutinina no plasma; e pessoas do gripo O não tem aglutinogênios na hemácias, mas possuem as duas aglutininas, anti-A e anti-B, no plasma.
Veja na tabela abaixo a compatibilidade entre os diversos tipos de sangue:

Tipos possíveis de transfusão
As aglutinações que caracterizam as incompatibilidades sanguíneas do sistema acontecem quando uma pessoa possuidora de determinada aglutinina recebe sangue com o aglutinogênio correspondente.
Indivíduos do grupo A não podem doar sangue para indivíduos do grupo B, porque as hemácias A, ao entrarem na corrente sanguínea do receptor B, são imediatamente aglutinadas pelo anti-A nele presente. A recíproca é verdadeira: indivíduos do grupo B não podem doar sangue para indivíduos do grupo A. Tampouco indivíduos A, B ou AB podem doar sangue para indivíduos O, uma vez que estes têm aglutininas anti-A e anti-B, que aglutinam as hemácias portadoras de aglutinogênios A e B ou de ambos.
Assim, o aspecto realmente importante da transfusão é o tipo de aglutinogênio da hemácia do doador e o tipo de aglutinina do plasma do receptor. Indivíduos do tipo O podem doar sangue para qualquer pessoa, porque não possuem aglutinogênios A e B em suas hemácias. Indivíduos, AB, por outro lado, podem receber qualquer tipo de sangue, porque não possuem aglutininas no plasma. Por isso, indivíduos do grupo O são chamadas de doadores universais, enquanto os do tipo AB são receptores universais.
|  |
Como ocorre a Herança dos Grupos Sanguíneos no Sistema ABO?
A produção de aglutinogênios A e B são determinadas, respectivamente, pelos genes I A e I B. Um terceiro gene, chamado i, condiciona a não produção de aglutinogênios. Trata-se, portanto de um caso de alelos múltiplos. Entre os genes I A e I B há co-dominância (I A = I B), mas cada um deles domina o gene i (I A > i e I B> i).

A partir desses conhecimentos fica claro que se uma pessoa do tipo sanguíneo A recebe sangue tipo B as hemácias contidas no sangue doado seriam aglutinadas pelas aglutininas anti-B do receptor e vice-versa.
Ausência de dominância
Ausência de dominância

Mas existem casos em que um gene interage com seu alelo, de maneira que o hibrido ou heterozigoto apresenta um fenótipo diferente e intermediário em relação aos pais homozigotos ou então expressa simultaneamente os dois fenótipos paternos. Fala-se então, emausência de dominância.
Podemos identificar dois tipos básicos de ausência de dominância, cujos estudos foram desenvolvidos em épocas posteriores à de Mendel: a herança intermediária e a co-dominância.

A herança intermediária é o tipo de dominância em que o indivíduo heterozigoto exibe um fenótipo diferente e intermediário em relação aos genitores homozigotos.Vejamos os seguintes exemplos:
Exemplo 1. A planta "maravilha" (Mirabilis jalapa) apresenta duas variedades básicas para a coloração das flores: a variedade alba(com flores brancas) e a variedade rubra (com flores vermelhas). chamando o gene que condiciona flores brancas de B e o gene para flores vermelhas de V, o genótipo de uma planta com flores brancas é BB, e o genótipo de uma planta com flores rubras é VV. Cruzando-se esses dois tipos de plantas (VV X BB), os descendentes serão todos VB; as flores dessas plantas (VB) serão rosas, isto é, exibirão um fenótipo intermediário em relação aos fenótipos paternais(flores vermelhas e brancas).
Exemplo 2. Nas galinhas de raça andaluza, o cruzamento de um galo de plumagem preta(PP) com uma galinha de plumagem branca(BB) produz descendentes com plumagem azulada (PB). Percebe-se então que a interação do gene para a plumagem preta(P) com o gene para plumagem branca(B) determina o surgimento de um fenótipo intermediário(plumagem azulada).
Co-dominância

A co-dominância é o tipo de ausência de dominância em que o indivíduo heterozigoto expressa simultaneamente os dois fenótipos paternos. Como exemplo podemos considerar da cor da pelagem em bovinos da raça Shorthon: os indivíduos homozigotos AA tem pelagem vermelha; os homozigotos BB tem pelagem branca;e os heterozigotos AB têm pêlos brancos e pêlos vermelhos alternadamente distribuídos.
quarta-feira, 27 de setembro de 2017
Primeira Lei de Mendel
Primeira Lei de Mendel
A primeira lei de Mendel nos diz que os descendentes recebem um alelo do par de genes que determina uma característica
Gregor Johan Mendel foi um monge agostiniano nascido no ano de 1822 que se interessou em explicar como as características dos pais são transmitidas a seus descendentes. Conhecido como o pai da genética, Mendel realizou todas as suas pesquisas sobre hereditariedade com ervilhas de cheiro (Pisum sativa), escolha que foi uma das razões de seu sucesso com suas pesquisas, pois essa leguminosa apresenta diversas vantagens como fácil cultivo, produção de grande quantidade de sementes, ciclo de vida curto, além de características contrastantes e de fácil identificação. Outro fato que contribuiu para o sucesso das pesquisas de Mendel foi que ele analisou apenas uma característica de cada vez, sem se preocupar com as demais características.
Em seus experimentos, Mendel teve o cuidado de utilizar apenas plantas de linhagens puras, por exemplo, plantas de sementes verdes que só originassem sementes verdes e plantas de sementes amarelas que só originassem sementes amarelas. Você deve estar se perguntando, como Mendel sabia que as plantas eram puras? Pois bem, para que ele tivesse certeza de qual planta era pura, ele as observava durante seis gerações, período de aproximadamente dois anos. Se durante essas gerações as plantas originassem indivíduos diferentes da planta inicial, elas não eram consideradas puras, mas se ocorresse o contrário e elas só originassem descendentes com as mesmas características da planta inicial, eram consideradas puras.

Uma vez constatado que as plantas eram puras, Mendel escolheu uma característica, por exemplo, plantas puras de sementes amarelas com plantas puras de sementes verdes, e realizou o cruzamento. Essa primeira geração foi chamada de geração parental ou geração P. Como resultado desse cruzamento, Mendel obteve todas as sementes de cor amarela e a essa geração denominou de geração F1. Os indivíduos obtidos nesse cruzamento foram chamados por Mendel de híbridos, pois eles descendiam de pais com características diferentes.
Em seguida, Mendel realizou uma autofecundação entre os indivíduos da geração F1, chamando essa segunda geração de geração F2. Como resultado dessa autofecundação, Mendel obteve três sementes amarelas e uma semente verde (3:1). A partir dos resultados obtidos, Mendel concluiu que como a cor verde não apareceu na geração F1, mas reapareceu na geração F2, as sementes verdes tinham um fator que era recessivo, enquanto as sementes amarelas tinham um fator dominante. Por esse motivo, Mendel chamou as sementes verdes de recessivas e as sementes amarelas de dominantes.
Em diversos outros experimentos, Mendel observou características diferentes na planta, como altura da planta, cor da flor, cor da casca da semente, e notou que em todas elas algumas características sempre se sobressaíam às outras.
Diante desses resultados, Mendel pôde concluir que:
→ Cada ser vivo é único e possui um par de genes para cada característica;
→ As características hereditárias são herdadas metade do pai e metade da mãe;
→ Os genes são transmitidos através dos genes;
→ Os descendentes herdarão apenas um gene de cada característica de seus pais, ou seja, para uma determinada característica, haverá apenas um gene do par, tanto da mãe quanto do pai.
Dessa forma, podemos enunciar a primeira lei de Mendel, também chamada de lei da segregação dos fatores da seguinte forma: “Todas as características de um indivíduo são determinadas por genes que se segregam, separam-se, durante a formação dos gametas, sendo que, assim, pai e mãe transmitem apenas um gene para seus descendentes”.
Segunda Lei de Mendel
Segunda Lei de Mendel

A segunda lei de Mendel ou também enunciada por diibridismo, refere-se à segregação independente dos fatores, isto é, a separação de dois ou mais pares de genes alelos localizados em diferentes pares de cromossomos homólogos, para formação dos gametas.
O princípio para essa segregação tem suporte na anáfase I da divisão meiótica, instante em que ocorre o afastamento dos cromossomos homólogos (duplicados), paralelamente dispostos ao longo do fuso meiótico celular.
Dessa forma, a proposição da segunda lei de Mendel, tem como fundamento a análise dos resultados decorrentes às possibilidades que envolvem não mais o estudo de uma característica isolada (Primeira Lei de Mendel), mas o comportamento fenotípico envolvendo duas ou mais características, em conseqüência da probabilidade (combinação) de agrupamentos distintos quanto à separação dos fatores (genes alelos / genótipo) na formação dos gametas.
Segue abaixo um exemplo prático da Segunda lei de Mendel:
Do cruzamento de ervilhas com características puras, em homozigose dominante e recessiva respectivamente para a cor da semente (amarela e verde) e para a textura da semente (lisa e rugosa), temos a seguinte representação para a geração parental e seus gametas:
RRVV (semente lisa e amarela) x rrvv (semente rugosa e verde)
Gameta → RV Gameta → rv
Desse cruzamento são originados exemplares vegetais de ervilha 100% heterozigóticas RrVv, com característica essencialmente lisa e amarela (geração F1 – primeira geração filial).
A partir do cruzamento entre organismos da geração F1, são formados tipos diferentes de gametas e combinações diversas para constituição dos indivíduos que irão surgir após a fecundação (geração F2).
Tipos de gametas da geração F1 → RV, Rv, rV e rv
Prováveis combinações entre os gametas:
9/16 → ervilhas com característica lisa e amarela;
3/16 → ervilhas com característica lisa e verde;
3/16 → ervilhas com característica rugosa e amarela;
1/16 → ervilhas com característica rugosa e verde.
Mendel concluiu que as características analisadas não dependiam uma das outras, portanto, são consideradas características independentes.
Nenhum comentário:
Postar um comentário